
Effects Of Prescribed Fire In Mixed Conifer Forest, Crater Lake National Park, Oregon by Ronald J. and Joy D. Mastrogiuseppe

Note: Ron Mastrogiuseppe was the founder of Crater Lake Institute
Complete PDF file prescribed-fire-mixed-conifer
Abstract
An investigation of fire effects was conducted from prescribed underburning in mixed conifer forest within a 973 acre area of Crater Lake National Park (CLNP) called the Panhandle. The primary objective of prescribed fire management was to restore and perpetuate the natural role of fire in natural areas of the park. Specific attributes of the mixed conifer forest which were investigated were preburn and postburn tree densities, understory vegetation response, and conifer seedling establishment and survival. Before burning, the study area was characterized by an abundant understory of white fir (Abies concolor (Gord. and Glend.) Lindl.) trees largely established since 1900. Some managers believe that this represents an unnatural abundance of A. concolor, primarily due to fire exclusion since the park’s establishment in 1902. Preburn densities of understory A. concolor of all size classes were estimated to be 4,960 trees/ha (2008/ac). Densities for all size classes ten years after the burn were estimated to be 3,354 trees/ha (1358/ac). Preburn densities of A. concolor > 1 dm dbh were estimated to be 4,240 trees/ha (1717/ac), while postburn densities were estimated to be 363 trees/ha (147/ac), a reduction in density of over 90 percent during one decade. Factors affecting mortality of A. concolor and the overstory ponderosa pine (Pinus ponderosa Dougl.) and sugar pine (P. Iambertiana Dougl.) included cambial and foliage scorch, disease, and insect infestations. During the ten years following the burn, mortality of the overstory P. ponderosa (> 5 dm dbh) has been at least 25 trees per ha (10/ac). Mortality has been brought about by attacks of the mountain pine beetle (Dendroctonus ponderosae Hopk.), and the western pine beetle (D. brevicomis). Local areas within the CLNP Panhandle forest are now distant from remaining seed sources for the two key pine species, and seedling recruitment during the postfire decade has been dominated by A. concolor. The survival ratio of conifer seedlings in the study area has been five A. concolor seedlings to one P. ponderosa seedling.
Introduction
The implementation of a fire management program which includes the prescribed use of fire requires specific information on the historical role of fire in the various ecosystems to be managed. Historical documents may contain information on the structure and composition of the presettlement vegetation. The goal of reconstructing historic processes and events results in a better understanding of present vegetation structure and composition (Bonnicksen and Stone 1981, 1985). Review of the fire history records for the area around Crater Lake, Oregon, provides some information on historical fire frequencies and intensities. Historical evidence exists for stand-replacing fires in Crater Lake National Park (CLNP) conifer forests dominated by lodgepole pine (Pinus contorta ssp. murrayana (Balf.) Critchfield), Pacific ponderosa pine (P. ponderosa Dougl.), noble fir (Abies procera Rehder), and mountain hemlock (Tsuga mertensiana (Bong.) Carr.) (Leiberg 1900, Langville et al. 1903). Frequently, photographs which accompany these historical descriptions display standing firekilled trees with a shrub understory and represent the initial sere of vegetation recovery. The Middle Fork of the Rogue River, near CLNP, has been the scene of two crown fires: one about 1880 and the other in 1910 (USDA For. Ser. 1916). An old newspaper account mentions fires within the then-proposed park boundary by emphasizing, “…5,000 acres of forest which have been absolutely destroyed by fire” (The Oregonian, 24 February, 1898). Although the location of this burn is not given in the newspaper, there are three possibilities: the Upper Middle Fork of Anna [Annie] Creek Burn, the Grayback Ridge Burn, or the West Road Burn. All these burns are largely in forests dominated by A. procera. Coville (1898) also reported a destructive fire of over 6100 ha (15,000 ac) east of Crater Lake.
During the late 1890s, fire control activities were initiated on national forest reserves adjacent to the park, with varying degrees of success (Brown 1960). Large numbers of blackened old-growth snags and stump remnants provide testimony to the crown fires which swept through mature stands during this period (USDA For. Ser. 1916). Fire suppression efficiency improved significantly during the 1930s, beginning a 50-year minimum of organized fire exclusion. The significance of the fire exclusion period in terms of vegetation change needs to be interpreted. For vegetation similar to the mixed-conifer forest of CLNP, mean fire return intervals (MFls) range from 11 to 24 years for six 16/ha (40/ac) plots (Bork 1984). One community within the Panhandle, dominated by A. concolor end identified as the pipsissewa-pyrola (Chimaphila umbellata-Pyrola picta) community (McNeil 1975), has not burned for over 130 years (last fire date being 1846). Shorter MFls reported for Pinus ponderosa-dominated habitat types of central Oregon include 17 years for Watkins Butte (Keen 1937) and 12 years for Black Butte (Sherman 1969). For ponderosa pine-grand fir (P. ponderosa-Abies grandis) habitat types reported MFls range from 25 to 30 years (Soeriaatmadja 1966).
One investigation of fire history within the study area reported fire dates ranging from 1750 to 1902, the year of park establishment (McNeil and Zobel 1980). The MFI reported ranged from 9 to 42 years. This investigation found no correspondence between fires burning east and west of the Annie Creek Canyon and no correspondence between fire years and the drought years reported by Keen (1937).
The Crater Lake Fire Management Plan was implemented during the 1976 fire season within the eastern portion of the Panhandle east of Annie Creek and east of State Highway 62. The burn objective was to reduce dead and down fuels at least 50 percent and to reduce the density of understory A. concolor. Dramatic increases in the density of A. concolor have sparked numerous investigations of the presettlement role of fire. The development of a shade-tolerant understory has been attributed by many investigators to fire exclusion (Merkle 1955, Kilgore and Taylor 1979, Bonnicksen and Stone 1981). Succession to A. concolor in giant sequoia- mixed conifer forests is closely linked to fire exclusion (Harvey, et. al. 1980), which led to the gradual elimination of tree aggregations characteristic of presettlement forests (Bonnicksen and Stone 1981, 1985).
The objectives of this study were to determine the effects of fire on (1) the density of the understory A. concolor established since 1900, (2) the associated understory vegetation response, and (3) conifer seedling establishment and survival.
Study Area
The study area is located within the southern part of the Panhandle area of Crater Lake National Park (CLNP), Klamath County, Oregon, along the east slope of the Cascade Mountain Range. Sample sites (elevation cat 1400 m) are both east and west of the narrow canyon of Annie Creek. The entire area was buried with pumice during the climactic eruption of ancient Mount Mazama 6840 yrs. B.P. or cat 7700 yrs. calibrated radiocarbon scale (Williams 1942, Bacon 1983). Soils developed from Mazama pumice resemble the Lapine series (Dyrness and Youngberg 1966) and the Steiger series (U.S. Bureau of Indian Affairs, 1958). The high porosity of these deep pumice deposits allows a high water-retention capacity. Mean precipitation ranges from 600 to 900 mm per year with 70 percent falling between November and May, primarily as snow (Sternes 1963). The first American settlements in the study area date from the early 1860s, when the park area was first traversed by a wagon road. The park was established in 1902, and the south Panhandle was added in 1932 as an entrance corridor. The Panhandle addition was made for the aesthetic value of the many magnificent old-growth Pinus ponderosa and P. Iambertiana beautifully displayed in this area.
Methods
Twelve permanent macroplots (25 X 15 m, 375 m2) were located within the proposed burn areas and marked with re-bar, enabling future inventories to monitor vegetation change. Control plots were located in the unburned area. Methods of data collection within these plots were similar to those of Daubenmire and Daubenmire (1968). All trees were tallied according to height and diameter at breast height (dbh) classes. Seedlings (less than 20 cm tall) were counted. These counts were repeated ten years after the burn. Coverage for each shrub and herb species was estimated using 50 microplots (20 X 50 cm, 0.1 m2) per macroplot. Ages of selected A. concolor were determined with an increment borer at 20 cm above the ground. Twenty trees of each diameter class were cut at ground level near the study plots, and their ages were determined through ring counts in the cross-section. Photopoints were established near each macroplot to provide a photographic record of the vegetation before and after the burning treatment. Photopoints were also established along the east and west flanks of State Highway 62 to record forest profile changes resulting from burning treatments.
Dead and down fuels were sampled along two transects per macroplot according to the procedures of Brown (1974). Fire weather measurements and estimates of fire behavior were made during the actual burning treatments. Two burning treatments were implemented, during July and September, 1976. All postburn conifer seedlings appearing within one study plot (70 X 70 m) were inventoried and identified with a numbered stake. Seedling establishment and survival were tallied during a six-year period following burning, and again ten years postburn.
Results
At the time of the prescribed burn, fuel moistures for 10-hr time lag dead fuels ranged from 12 to 20 percent. Relative humidity varied from 40 to 60 percent, and wind within the stand was less than five mph. Fire behavior varied from local torching into the high canopy to creeping backfire with flame lengths from one to three ft. As a result of the fire, down and dead woody fuel loadings (2.5 to 3.0 tons/ha) were reduced 70 percent in the one-hr time lag class. In the 10-hr class, fuel loadings (5.5 to 6.8 tons/ha) were reduced 67 percent. However, fuels in the 100-hr and 1000-hr classes increased an estimated 25 percent (25 to 40 tons/ha) during the following five years, as a result of tree attrition. Fuel loadings (one-hr and 10-hr time lag) 10 years postburn, estimated at two to six tons/ha, are sufficient to carry another fire.
Preburn understory A. concolor densities of all size classes were estimated to be 4,960 trees/ha (Table 1). Of these, 78 percent were over one m in height and under two dm in diameter (dbh). Postburn densities for all size classes ten years after the burn were estimated to be 3354 trees/ha.
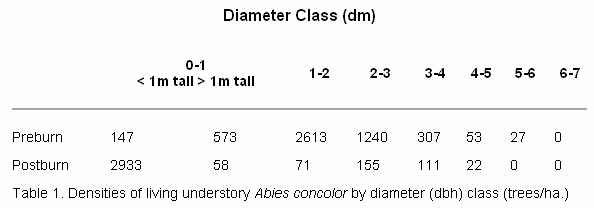
Preburn understory densities of trees over one dm dbh were estimated to be 4240 trees/ha. Densities of these larger trees one decade following the burn were estimated to be 363 trees/ha, a reduction of over 90 percent (Table 1). Many of the understory A. concolor which survived the actual fire exhibited basal bark cracking and peeling from cambial injury. These individuals died within two growing seasons after the burn.
The vegetation changes resulting from prescribed fire are not restricted to understory conifers. Significant numbers of overstory P. ponderosa were lost during the postfire decade as a result of insect attacks. Preburn densities of living P. ponderosa over one dm dbh were estimated to be 195 trees/ha. One decade following the burn, between 12 and 22 percent of P. ponderosa (all size classes) had died in sampled areas.
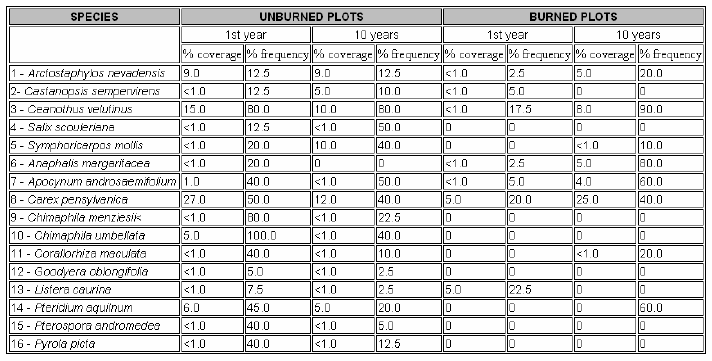
Following the fire, the first evidence of plant growth on the ashbed was Pyronema sp., a bright orange-colored fungus especially common in old-stump depressions. (Occasional stumps (one to three per ha) had been left following the selective cutting of insect-infested pines during the early years of park insect control programs.) Wherever canopy gaps were created through fire-induced mortality, shrub and herbaceous seedlings were observed. The drought conditions during late 1976 through 1977, however, may have been a significant factor affecting seedling establishment and survival. Understory shrub and herb species present and comprising early- to mid-postfire succession are listed in Table 2.
The study area seedling plot (70 X 70 m) carried the more intense fire, creating large canopy gaps. For this plot the following seedling estimates were made 10 years postburn: A. concolor > 5200/ha, P. ponderosa > 930/ha, P. contorta subsp. murrayana > 110/ha, and P. Iambertiana > 7/ha. Among the seedlings established in 1976-77, the tallest (> 1.5 m) are P. contorta subsp. murrayana.
Discussion
Dominant trees. This investigation demonstrates that significant numbers of understory A. concolor can be removed through fire mortality and subsequent mortality induced by disease and insects (Table 1). Natural fire creates and maintains mosaics of different ages and species since these fires burn during periods of variable fire weather conditions with different intensities and effects. It is believed that the role of natural fire was to perpetuate a fire disclimax of P. ponderosa. In the ecotonal areas of the CLNP Panhandle, however, both ponderosa pine and white fir have been favored during the first phase of prescription burning. More recent intense burning during the 1980s (in areas west of State Highway 62) increased the mortality of dominant old-growth pines.
The general distribution of A. concolor has not changed significantly since the pre-fire suppression period (Leiberg 1900, White Fir Distribution Map, Pl. LXXX). One early description (Kerr 1913) of A. concolor-P. ponderosa stands is noteworthy:
“Yellow pine and White fir are numerically the most important species and vary in their relative proportions from pure Yellow pine on some of the lower slopes and flats to the pure fir type of the upper slopes where White fir constitutes over 50% of the volume. On the drier sites, White fir commonly occurs in pure clumps scattered through the Yellow pine stand…For the region as a whole, White fir comprises probably 25% of the total stand….”
Early observations by Kerr (1913) of fire behavior in similar stands provide one interpretation of fire effects:
“White fir seedlings that are exposed to fire are usually badly injured, but they possess an exceptionally high degree of vitality, and unless the stem is completely girdled will ordinarily recover…Because of the unprotected resinous stems of Yellow pine seedlings, and the dry surface conditions of the sites on which they habitually grow, the lightest surface fire will ordinarily kill all young seedlings of this species. On the other hand, the dense cover of White fir clumps keeps the ground beneath cool and moist at all times, so that a light fire will not run under them, but is stopped at the edge with perhaps only a slight damage to the seedlings on the outer border. Scattered individuals would probably be killed, but since in the mixed stands, the majority of the White fir occurs in clumps, it follows that the damage to such a stand resulting from surface fire is very largely borne by the Yellow pine, and that repeated surface fires must cause a decrease in the proportion of Yellow pine in the stand.”
This interpretation, although contrary to that understood for many habitats dominated by P. ponderosa, may be applicable in broad ecotonal areas. Vegetation changes resulting from dynamic environmental variables, including climate and fire, are not unique to the fire suppression period, and have been described in several vegetation history studies (P. J. Mehringer, Jr., pers. comm. 1986). One key attribute of A. concolor is its shade tolerance (Minore 1979). Potential competing conifers are restricted by low levels of sunlight and a thick litter/duff layer which retains moisture. In addition, branch and foliage retention in the lower crown promotes torching and crowning fire behavior at low live fuel moistures. This may be especially detrimental to neighboring pines and the future pine seed source.
If another fire event occurs in the study area, seedling mortality is expected to be high, and the opportunity for additional P. ponderosa and P. lambertiana seedling establishment will be limited. Local areas within the Panhandle have already been changed through loss of P. ponderosa and P. lambertiana seed sources in terms of their immediate dispersal distance.
Understory Species Response. Fire response predictions can be based upon a knowledge of a species’ life history attributes characteristic of the reproductive and regenerative phases. Resilient preburn species typical of midsuccession indicate that the tolerance’ model of succession (Cornell and Slatyer 1977) is appropriate. Many of the plant species appearing soon after a fire are those already present in the prefire stands. They reproduce either from buried seeds or vegetatively from propagules. Sprouts of snowbrush (Ceanothus velutinus Dougl.) appeared during the 1977 growing season but showed evidence of browsing by deer or elk. Seed germination in this species is dependent upon fire and moisture to crack the seed coat. Ceanothus velutinus is important in nitrogen fixation, and its seedlings and sprouts may provide suitable microsites for the establishment of conifer seedlings (Zavitkovski and Newton 1968). However, vigorous shrubs of this species provide intense competition to young conifers.
Accumulations and decomposition of P. ponderosa litter may be detrimental to some herbaceous species (Moir 1966). Once the litter has been consumed by fire, the resulting seedbed should enhance the diversity of herbaceous species. However, the potential understory flora available to invade burned sites is depauperate, and this factor accounts for much openness in the forest understory. The only two herbaceous species new in the postburn community were thistle (Cirsium vulgare (Savi) Tenore) and pearly everlasting (Anaphalis margaritacea (L.) B. & H.) (Table 2). No new shrub species arrived following the burn.
Summary
Natural fire has been a significant ecological process within the P. ponderosa-A. concolor forests typical of lower slopes along the eastern Cascades. The objective to restore and perpetuate the role of natural fire within this mixed conifer forest is a goal which may be achieved through a better understanding of the effects of prescribed fire and additional fire history studies.
Natural area management objectives need to be measurable as well as unambiguous. Ecological information is essential in formulating these objectives. The primary objective at CLNP is to restore ecosystems to the condition which would have existed today without the interference with natural processes brought about by European settlement. Since vegetation is dynamic through time, resource managers are managing change in their efforts to perpetuate more natural assemblages of plant communities.
The dramatic increase in shade-tolerant, understory species during this century is generally interpreted to be the primary result of effective fire suppression activities. Fire suppression may be appropriate as only one factor in the increase of understory A. concolor within the Panhandle of Crater Lake National Park. Although fire and fire-induced decay may be effective in removing large numbers of understory conifers during several years, new arrivals from the abundant seed of A. concolor in the high canopy must be considered. In the CLNP Panhandle study the survival ratio of A. concolor to P. ponderosa seedlings was five to one.
Disease, insect infestations, and fine root mortality resulting from intense fires have all reduced the seed source from dominant pines in many areas. Therefore, additional prescribed burning is not recommended at this time. Time without fire is essential for monitoring change resulting from these more intense fires and to allow the established pine regeneration to survive to a larger size. Many understory species are resilient, and open areas within canopy gaps will have a more diverse flora. Downed dead fuel loadings will continue to increase as standing dead trees disintegrate. To achieve more acceptable fuel loadings for these showcase areas which are highly visible in the Panhandle corridor, intensive manual labor is necessary to establish small piles of woody debris. These woody debris piles may be burned once sufficient snow cover occurs, to protect established cohorts of pine regeneration. Since fuel loadings will remain high, it is important that effective fire suppression be implemented should any lightning- or man-caused fires occur in the Panhandle. Once fuel loadings have been reduced to a more manageable level, lightning fires burning under prescription as natural fires will determine the nature of the future fire regime for this forest. Containment or suppression may be necessary for some of these future management fires, to achieve visitor safety and to perpetuate the critical pine component of the Panhandle forest. However, in the long term, to perpetuate a more natural forest ecosystem, low-intensity prescribed fire must be implemented to perpetuate the unique ecotonal mixed-conifer forest of the Panhandle. The two small control areas in the Panhandle should remain undisturbed and untreated as an example of forest vegetation prior to any manipulation.
Acknowledgements
This research project was supported in part by The National Park Service, Crater Lake National Park, Oregon. Among the park staff who provided assistance were: M. Forbes, G. Johannsen, S. Mark, D. Sholly, L. Smith, H. Tanski, and J. White. Their assistance and patience are greatly appreciated.
Literature Cited
Bacon, C.R. 1983. Eruptive history of Mount Mazama and Crater Lake Caldera, Cascade Range, U.S.A. In S. Aramaki and 1. Kushiro (Editors), Arc Volcanism. J. Volcanol. Geotherm. Res. 18:57-115. Bonnicksen, T.M., and E.C. Stone. 1981. The giant sequoia-mixed conifer forest community characterized through pattern analysis as a mosaic of aggregations. For. Ecology. and Manage. 3:307-328.
Bonnicksen, T.M., and E.C. Stone. 1985. Restoring naturalness to national parks. Envir. Manage. 9:479-486.
Bork, J.L. 1984. Fire history in three vegetation types on the eastern side of the Oregon Cascades. Oregon State University, Corvallis. Ph.D. Thesis.
Brown, C.E. 1960. History of the Rogue River National Forest, Oregon. Vol. I. 1893-1932. USDA For. Ser., Medford, Ore.
Brown, J.K. 1974. Handbook for inventorying downed woody material. USDA For. Ser. Gen. Tech. Rep. INT-16.
Connell, J.H., and R.O. Slatyer. 1977. Mechanisms of succession in natural communities and their role in community stability and organization. Amer. Nat. 111: 1119-1 144.
Coville, F. 1898. Forest growth and sheep grazing in the Cascade Mountains of Oregon. USDA Div. For. Bull. 15.
Daubenmire, R., and J. Daubenmire. 1968. Forest vegetation of eastern Washington and northern Idaho. Wash. Agr. Exp. Sta. Tech. Bull. 60. Washington State University. Pullman.
Dyrness, C.T., and C.T. Youngberg. 1958. Soil-vegetation relationships in the central Oregon pumice region. In First North American Forest Soils Conference. Mich. State Univ. Agr. Exp. Sta., East Lansing. Pp. 57-66.
Harvey, H.T., H.S. Shellhammer, and R.E. Stecker. 1980. Giant sequoia ecology, fire and reproduction. Sci. Monogr. Series No. 12. USDI, Park Ser.
Keen, F.P. 1937. Climatic cycles in eastern Oregon as indicated by tree rings. Monthly Weather Rev. 65:175-188.
Kerr, A.F. 1913. White fir in the Klamath Basin. For. Quart. 11:349-362.
Kilgore, B.M. and D. Taylor. 1979. Fire history of a sequoia-mixed conifer forest. Ecology 60:129-142.
Langville, H.D., F.G. Plummer, A. Dodwell, T.F. Roxon, and J.B. Leiberg. 1903. Forest conditions in the Cascade Range Forest Reserve, Oregon. USDI Geological Survey Prof. Paper No. 9. Washington, D.C.
Leiberg, J.B. 1900. The Cascade Range and Ashland Forest Reserves and adjacent regions. USDI Geological Survey 9 (In Twenty-First Annual Rep. of the Survey 1899-1900, Part V, Forest Reserves).
McNeil, R.C. 1975. Vegetation and fire history of a ponderosa pine-white fir forest in Crater Lake National Park. Oregon State University, Corvallis. M.S. Thesis.
McNeil, R.C., and D.B. Zobel. 1980. Vegetation and fire history of a ponderosa pine-white fir forest in Crater Lake National Park. Northw. Sci. 54:30-46.
Merkle, J. 1955. Abies concolor (white fir): A growing problem in the West. Abstr. in Bull. Ecology. Soc. Amer. 36:80.
Minore, D. 1979. Comparative autecological characteristics of northwestern tree species. A Literature Review. USDA For. Ser. Gen. Tech. Rep. PNW-87.
Moir, W.H. 1966. Influence of ponderosa pine on herbaceous vegetation. Ecology 47:1045-1048.
Sherman, R.J. 1969. Spatial and developmental patterns of the vegetation of Black Butte, Oregon. Oregon State University, Corvallis. Ph.D. Dissertation.
Soeriaatmadja, R.E. 1966. Fire history of the ponderosa pine forests of the Warm Springs Indian Reservation, Oregon. Oregon State University, Corvallis. Ph.D. Dissertation.
Sternes, G.L. 1963. Climate of Crater Lake National Park. Crater Lake Natural History Assoc. and USDI Park Ser.
USDA Forest Service. 1916. Fire History Report, Prospect District No. 1. Crater National Forest.
Volland, L.A. 1963. Phytosociology of the ponderosa pine type on pumice soils in the Upper Williamson River Basin, Klamath County, Oregon. Oregon State University, Corvallis. M.S. Thesis.
Williams, H. 1942. The geology of Crater Lake National Park, Oregon. Carnegie Inst. Wash. Publ. 540. Wash., D.C.
Zavitkovski, J., and M. Newton. 1968. Ecological importance of snowbrush, Ceanothus velutinus, in the Oregon Cascades. Ecology 49:1134-1145.
Other pages in this section
*** previous title *** --- *** next title ***